
 資料結構
資料結構 網路
網路 關係型資料庫管理系統
關係型資料庫管理系統 作業系統
作業系統 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 程式設計
C 程式設計 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP蛋白質靶向或蛋白質分選
蛋白質靶向
蛋白質被傳遞到細胞內或細胞外的適當位置的生物學過程稱為蛋白質靶向或蛋白質分選。蛋白質可以分泌到細胞外部、質膜、各種細胞內膜、細胞器的內部或質膜。
蛋白質本身包含控制傳遞機制的資訊。正確分選對細胞至關重要,分選出現問題與許多疾病有關。
簡介
在真核細胞中,各種蛋白質必須被傳遞到細胞內的不同位置,或者在某些情況下,被輸出到細胞外環境。適當的蛋白質是如何到達相應位置的?
蛋白質透過細胞中存在的各種運輸機制被傳遞到其目標位置,有點像分子郵政服務。在這些系統中,蛋白質使用分子標籤(通常是氨基酸序列)進行“地址”標記,以便傳遞到特定的位置。讓我們看看這些傳遞系統是如何工作的。
訊號肽
訊號肽透過充當靶向訊號,使細胞運輸系統能夠將蛋白質引導到特定的細胞內或細胞外區域。儘管尚未發現訊號肽的共有序列,但其中許多仍然具有獨特的結構三部分:
靠近N端的親水性、帶正電的區域。
訊號肽中一段 10 到 15 個疏水性氨基酸的區域。
靠近C端的略微極性區域,當它們位於靠近切割點時,有利於具有較短側鏈的氨基酸。
蛋白質到達目的地後,訊號肽通常會被訊號肽酶切割。
因此,大多數成熟蛋白質中不存在訊號肽。雖然過氧化物酶體的C端延伸部分存在靶向序列,但大多數訊號肽位於N端。
蛋白質易位
用於分泌或特定細胞器的蛋白質必須被運輸,因為核糖體將mRNA翻譯成蛋白質是在細胞質中進行的。這個過程被稱為共翻譯易位,它也可以在翻譯過程中發生,或者翻譯後易位,它發生在翻譯完成後。
蛋白質靶向
線粒體
大多數線粒體蛋白作為具有攝取肽訊號的胞質前體產生,而細胞器中的一些蛋白質是由線粒體DNA產生的。
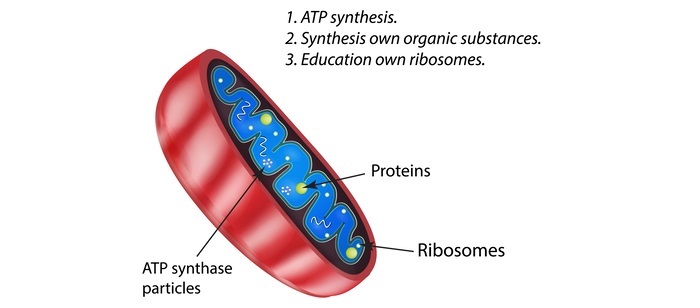
根據其序列,由胞質伴侶蛋白hsp70結合併發送到線粒體的未摺疊蛋白可能定位於四個不同的位置。它們可能被引導到內膜、膜間隙、外膜或線粒體基質。健康和疾病與其中一種或多種機制的缺陷有關。
線粒體基質
當多肽到達基質時,訊號序列會被加工肽酶切割,剩餘的序列在等待適當摺疊和作用時會被線粒體伴侶蛋白保持。
線粒體在氧化磷酸化過程中產生的電化學梯度推動並拉動多肽從胞質到膜間隙,然後到基質。
由於積極參與代謝的線粒體,膜間隙具有正電位,而基質具有負電位。靶向序列的帶正電部分被基質的負電位引導到其預期位置。
線粒體內膜
根據其整體序列,線粒體蛋白可以透過三種不同的途徑之一被引導到內膜,儘管它們仍然可以透過使用TOM20/22匯入受體複合物和TOM40通用匯入核心從外膜進入。
在靶向內膜的蛋白質的第一條途徑中,其具有將多肽引導到包含先前所述易位複合物的內膜複合物的基質靶向序列,其步驟與靶向基質的蛋白質的步驟相同。
線粒體外膜
前體蛋白透過使用內部靶向序列產生跨磷脂雙層的疏水性α螺旋或β桶,與外膜易位複合物相互作用以將其嵌入膜中。此過程稱為外膜靶向。
這可能透過兩種不同的途徑發生,具體取決於前蛋白的內在序列。如果前蛋白具有能夠構建α螺旋的內部疏水區域,則它將使用線粒體匯入複合物並橫向運輸到膜中。
與β桶構建蛋白相關的並且具有疏水性內部序列的前蛋白將從上述外膜複合物TOM20/22匯入到膜間隙。
當它們與TIM9/10膜間隙蛋白複合物結合時,它們會被轉移到外膜的分選和組裝機制(SAM),該機制將靶向蛋白作為β桶橫向化。
葉綠體
根據其序列,蛋白質可以被髮送到葉綠體的基質、類囊體腔、外被膜、內被膜或類囊體膜。
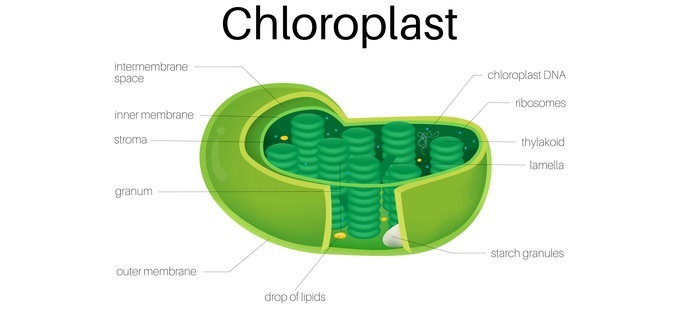
由於它們通常缺乏可切割的分選序列,因此靶向葉綠體被膜的蛋白質會被膜分選複合物橫向置換。
葉綠體被膜內的Toc和Tic複合物對於從胞質中大量匯入前蛋白是必需的。
葉綠體外部膜的易位酶稱為Toc,葉綠體內部膜的易位酶稱為Tic。Toc複合物的功能由至少三種蛋白質組成。
基質
Tic複合物中必須存在至少五種不同的Tic蛋白才能形成穿過內被膜的易位通道。到達基質後,基質匯入序列會被訊號肽酶切除。
根據目前的知識,ATP透過基質HSP伴侶蛋白的水解驅動這種蛋白質運輸機制到基質,而不是線上粒體中產生的推動蛋白質匯入的跨膜電化學梯度。
結論
雙靶向肽通常具有介於兩個特定肽之間的特徵。這些蛋白質的靶向肽包含相對較少的帶負電荷的氨基酸和大量的鹼性和疏水性氨基酸。
它們包含較少的丙氨酸和更多的亮氨酸和苯丙氨酸。與線粒體和葉綠體蛋白相比,雙靶向蛋白具有更疏水的靶向肽。僅根據其物理化學特性來確定肽的雙靶向狀態是費力的。

2K+ 次瀏覽
