
 資料結構
資料結構 網路
網路 關係型資料庫管理系統 (RDBMS)
關係型資料庫管理系統 (RDBMS) 作業系統
作業系統 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C語言程式設計
C語言程式設計 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP訊號肽:概述
介紹
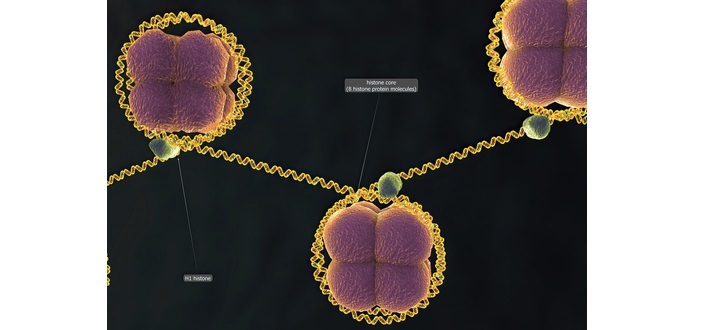
訊號肽,也稱為訊號序列,是一種短肽,通常長16到30個氨基酸。它也偶爾可以在大多數新合成蛋白的C端或內部區域非經典地找到,這些蛋白正走向分泌途徑。
這些蛋白質包括那些插入大多數細胞膜或存在於特定細胞器(如內質網、高爾基體或內體)中的蛋白質。大多數II型和多跨膜結合蛋白是由它們的第一個跨膜結構域導向分泌途徑的,該結構域在生化上類似於訊號序列,但不被切割。
雖然大多數I型膜結合蛋白具有訊號肽,但II型和多跨膜結合蛋白並非如此。
功能
訊號肽提示細胞易位蛋白質,通常到細胞膜。
原核生物中的訊號肽將新生成的蛋白質驅動到質膜的SecYEG蛋白傳導通道。
在真核生物中,存在類似的機制,其中訊號肽將新生成的蛋白質引導到位於內質網中的Sec61通道,該通道與SecYEG具有結構和序列相似性。
透過該通道(包括SecYEG和Sec61通道)的易位被稱為易位子。
跨膜結構域可以擴散透過易位子中的側向門,以分配到周圍膜中,而分泌蛋白則穿過通道。
結構
“h區”——構成訊號肽核心的大量疏水氨基酸片段,長度在5到16個殘基之間——傾向於盤繞成單個α螺旋。
此外,許多訊號肽以一段帶正電荷的短氨基酸序列開始,這可能支援“正電荷內側規則”在易位過程中維持多肽的正確結構。
由於其靠近N端,因此被稱為“n區”。訊號肽酶通常識別並切割訊號肽末端的氨基酸片段,這就是該區域被稱為切割位點的原因。
然而,作為訊號肽起作用的跨膜結構域(也稱為訊號錨定序列)缺乏這個切割位點。
為了產生成熟的蛋白質和遊離的訊號肽,訊號肽酶可能在易位完成之前或之後進行切割。然後,某些蛋白酶降解遊離的訊號肽。
此外,不同的訊號肽型別針對不同的目標區域。例如,針對線粒體環境的目標肽具有不同的長度,並且其結構中短的帶正電荷區域和疏水區域交替出現。
蛋白質的N端和C端都包含核定位訊號肽,這些訊號肽通常保留在成熟蛋白中。
共翻譯途徑 |
翻譯後途徑 |
|---|---|
訊號肽離開核糖體並被訊號識別顆粒(SRP)識別後,就會啟動共翻譯途徑。 |
當蛋白質合成完成後,就會啟動翻譯後過程。在原核生物中,SecB伴侶蛋白識別翻譯後底物的訊號序列,並將蛋白質轉移到SecA ATP酶,然後SecA ATP酶將蛋白質泵入易位子。 |
隨後,訊號序列-核糖體-mRNA複合物被引導到SRP受體,該受體位於質膜(在原核生物中)或內質網的表面,其中翻譯停滯只發生在真核生物中(在真核生物中)。 |
雖然已知真核生物中存在翻譯後易位,但對其瞭解甚少。 |
膜靶向後,訊號序列被引入易位子。當核糖體物理地附著在易位子的胞質面上時,蛋白質合成就會恢復。 |
已知易位子和另外兩種膜結合蛋白Sec62和Sec63對於酵母翻譯後易位是必需的。 |
訊號識別顆粒 (SRP)
只有內質網,而不是其他型別的膜,參與分泌蛋白和其他非胞質蛋白的產生。
因此,內質網膜必須具有獨特的特徵。一旦內質網訊號肽離開核糖體,由六個多肽鏈和一個300核苷酸RNA組成的SRP就會與其結合。
SRP受體
被稱為SRP受體的650個殘基整合膜蛋白介導核糖體/多肽/SRP複合物與膜的相互作用。它可以與SRP和核糖體結合。
這種蛋白質在粗麵內質網的胞質表面可見。仔細觀察發現,存在GTP結合結構域。
訊號肽酶
各種輔助因子使正確解釋分選訊號和將所有蛋白質輸送到其預期位置成為可能。訊號肽酶是負責從蛋白質中去除訊號肽序列的專門酶。
一旦蛋白質被訊號肽導向到所需位置,這些酶就會被啟用。大多數細胞器隔室都具有獨特的變體,這些變體只能切割一類成員,而不能切割其他成員。
結論
雖然兩者有時含糊地被稱為“前導肽”,但訊號肽不應與有時由前導mRNA編碼的前導肽混淆。
這些額外的前導肽是短多肽,不參與最終蛋白質序列的構成,它們可能控制主要蛋白質的轉錄或翻譯,而不是參與蛋白質定位。
雖然真核基因的調控採用類似的方法,稱為uORFs(上游開放閱讀框),但這種型別的引導肽通常與細菌中觀察到的一種基因調控形式有關。

瀏覽量:144
